Il Disturbo dello Spettro Autistico (ASD) è una condizione neuroevolutiva complessa che si manifesta attraverso sfide nella comunicazione, difficoltà nelle interazioni sociali e comportamenti ristretti e ripetitivi. La sua eziologia è multifattoriale, intrecciando una complessa architettura genetica con diverse caratteristiche morfologiche che contribuiscono alla sua eterogeneità clinica e comportamentale. La ricerca scientifica, attingendo a varie fonti tra cui Google Scholar, si impegna a decifrare questi meccanismi sottostanti per migliorare la comprensione, la diagnosi precoce e gli interventi terapeutici.
Introduzione al Disturbo dello Spettro Autistico
Il DSM-5 classifica l'ASD come un disturbo del neurosviluppo, caratterizzato da difficoltà significative nella comunicazione e nell'interazione sociale, unitamente a pattern di comportamento restrittivi e ripetitivi. La prima descrizione dell'autismo risale al 1943 ad opera del pediatra Leo Kanner, che studiò il disturbo seguendo 11 bambini. Da allora, gli esperti hanno dibattuto se l'ASD sia una condizione innata o acquisita. Per anni, la visione prevalente era che i fattori ambientali fossero i principali determinanti (Hyman et al., 2020). Successivamente, la ricerca si è spostata sulla genetica, indicando un forte contributo genetico, con tassi di concordanza più elevati tra i gemelli monozigoti (Hallmayer et al., 2011).
Le modifiche nei criteri diagnostici dell'ASD hanno unificato le diagnosi relative all'autismo, richiedendo ora una specifica combinazione di difficoltà nella socializzazione, nella comunicazione e nei comportamenti ripetitivi (Matson and Kozlowski, 2011). Questa crescita è attribuita principalmente ai cambiamenti nei criteri diagnostici e a una maggiore consapevolezza del disturbo (Thapar and Rutter, 2021). Inoltre, la ricerca di misure oggettive, come i biomarcatori, mira a supportare l'identificazione precoce dell'ASD, considerando la crescente necessità di diagnosi e interventi più tempestivi (Levin and Nelson, 2015).
La morfologia cerebrale nei bambini con autismo può includere caratteristiche quali un aumento del volume cerebrale, in particolare nell'emisfero sinistro, nonché variazioni nella materia grigia e bianca, riflettendo la complessità dello sviluppo neurale associato al disturbo. Questi aspetti sono essenziali per comprendere la base neurobiologica dell'autismo e le sue implicazioni per il comportamento e il funzionamento cognitivo (Lamanna e Meldolesi, 2024).
Nell'ambito di questo contesto, questo studio discute alcune alterazioni genetiche e morfologiche associate all'ASD, esplorando come questi cambiamenti possano contribuire alle alterazioni cerebrali osservate nel disturbo. Verrà inoltre affrontata la relazione tra queste alterazioni, con un focus su come le modifiche genetiche possano influenzare i cambiamenti cerebrali.
Caratteristiche Morfologiche nel Cervello degli Individui con ASD
Lo sviluppo cerebrale è un processo continuo che comporta significativi cambiamenti strutturali e funzionali. La comprensione di questi cambiamenti è essenziale per l'identificazione precoce di disturbi come l'ASD (Xie et al., 2023). Sebbene l'eterogeneità dell'autismo ponga una sfida clinica, le alterazioni cerebrali possono servire come biomarcatori per diagnosticare e comprendere questo complesso disturbo (Varcin e Nelson, 2016).
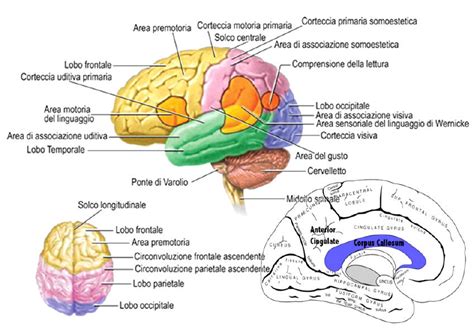
Gli individui con autismo spesso mostrano un eccessivo aumento del volume cerebrale nei primi anni di vita, seguito da un rallentamento nell'infanzia e, in alcuni casi, da un declino durante l'adolescenza e l'età adulta (Courchesne et al., 2011). In uno studio longitudinale di RM, i bambini con diagnosi di ASD hanno mostrato un aumento generalizzato del volume corticale a 2 anni di età. Inoltre, i bambini autistici hanno mostrato volumi significativamente maggiori di materia grigia e bianca. Gli autori dello studio hanno indicato che la crescita cerebrale eccessiva si verifica prevalentemente nei primi mesi dopo la nascita (Hazlett et al., 2011).
Studi precedenti hanno supportato l'ipotesi di un aumento del volume corticale nei bambini con autismo; tuttavia, non è stato osservato un corrispondente aumento del volume cerebellare. Inoltre, la circonferenza cranica di questi bambini appare normale alla nascita, ma inizia a crescere a un ritmo accelerato intorno ai 12 mesi di età (Hazlett et al., 2005).
Studi di RM dimostrano che, nonostante la crescita iniziale del volume cerebrale, si verifica un declino durante l'adolescenza e la prima età adulta (Roy e Uddin, 2021). Una ipotesi per spiegare questa crescita cerebrale eccessiva suggerisce che possa derivare da un maggior numero di neuroni e connessioni sinaptiche formate durante il periodo prenatale. Queste alterazioni atipiche dello sviluppo cerebrale sono considerate legate alle caratteristiche cliniche dell'ASD (Courchesne et al., 2011).
Peterson et al. (2021) suggeriscono che, sebbene ci sia un aumento iniziale del volume del liquido cerebrospinale extra-assiale (CSF) nei bambini autistici, questo volume tende a normalizzarsi nel tempo, specialmente dopo i 4 anni di età. Gli autori propongono che questa normalizzazione possa essere correlata alla maturazione cerebrale in corso durante l'adolescenza e l'età adulta.
In uno studio condotto da Rabelo et al. (2023), sono state identificate aree di disorganizzazione corticale nella corteccia prefrontale dorsolaterale (DL-PFC) di bambini con ASD e gruppi di controllo. Queste aree hanno mostrato un'espressione genica alterata, come CALB1, RORB e PCP4, in strati specifici, in particolare negli strati II-III. Queste aree hanno mostrato un rapporto glia-neurone (GNR) significativamente ridotto rispetto alle regioni non interessate e ai cervelli neurotipici. Inoltre, lo studio ha rivelato che il GNR nella DL-PFC dei bambini con ASD era, in media, inferiore del 20%, suggerendo una riduzione relativa delle cellule gliali o un aumento del numero di neuroni. Un aumento della proporzione di neuroni nella corteccia prefrontale degli individui con ASD è stato riportato anche in altri studi, come quello di Falcone et al. (2021) (Figura 1).

Stoner et al. (2014) hanno condotto un'analisi di campioni post-mortem di 22 cervelli, 11 dei quali appartenevano a bambini con ASD e 11 a controlli. Lo studio ha confrontato l'organizzazione degli strati corticali tra i gruppi. I risultati hanno rivelato che 10 dei 11 casi mostravano aree di disorganizzazione nella neocorteccia, indicando fallimenti nella migrazione neuronale durante lo sviluppo fetale. Queste anomalie suggeriscono che le alterazioni associate all'autismo possano originare in utero, influenzando la connettività cerebrale e le funzioni cognitive, sociali e linguistiche.
In uno studio parallelo, è stato analizzato l'impatto dei geni associati all'autismo sulla connettività tra diverse regioni corticali. È stato riscontrato che le alterazioni genetiche nell'ASD sono più pronunciate negli strati superficiali della corteccia. Inoltre, le mutazioni nei geni correlati all'ASD sono associate alla comunicazione interemisferica, che può compromettere l'integrazione delle informazioni provenienti da diverse aree del cervello (Parikshak et al., 2013).
Inoltre, uno studio di Mei et al. (2023), che ha utilizzato un'analisi multimodale di soggetti autistici e controlli, ha identificato alterazioni nel volume della materia grigia (GM) in varie regioni. È stato osservato un ridotto volume di GM in aree come l'insula, il giro frontale inferiore e la corteccia orbitofrontale, mentre è stato notato un aumento del volume in aree come la corteccia calcarina e il giro frontale medio. Gli autori hanno correlato queste alterazioni con il profilo comportamentale dell'ASD.
Questi risultati morfologici e funzionali sono sempre più compresi attraverso i progressi nella genetica. Infatti, la ricerca genetica ha rivelato come determinate mutazioni e varianti genetiche influenzino lo sviluppo e l'organizzazione del cervello, contribuendo alle anomalie osservate nella struttura e nella connettività cerebrale (Nalin et al., 2022).
Basi Genetiche dell'ASD
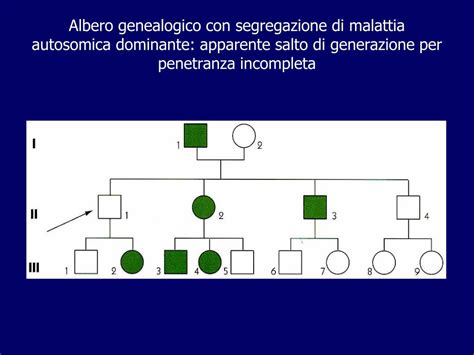
La relazione tra genetica e disturbi psichiatrici è complessa, e l'identificazione di geni individuali e la valutazione definitiva dei loro effetti rimangono sfide persistenti. Gli studi sui gemelli hanno contribuito a chiarire l'influenza genetica, mostrando che i gemelli identici hanno maggiori probabilità di ricevere una diagnosi di ASD rispetto ai gemelli fraterni (Colvert et al., 2015; Tick et al., 2016; Lichtenstein et al., 2010).
Da questa prospettiva, l'eziologia dell'ASD coinvolge una complessa interazione tra fattori genetici e ambientali, modulata da processi epigenetici (Genovese e Butler, 2023). Uno studio condotto da Bai et al. (2019) su 22.156 bambini in cinque paesi ha identificato che la genetica è responsabile di circa l'80% dei casi. Il rimodellamento della cromatina coinvolge cambiamenti nella struttura del DNA e degli istoni senza alterare la sequenza stessa, regolando l'espressione genica e influenzando la differenziazione neuronale e la formazione delle sinapsi (Genovese e Butler, 2023).
In questo contesto, l'ASD può essere classificato in due forme: sindromica e non sindromica. L'ASD sindromico si riferisce alla presenza di specifiche mutazioni genetiche che si manifestano come sindromi neurologiche riconoscibili, come la sindrome dell'X fragile, la sindrome di Rett e la sclerosi tuberosa (Nóbrega et al., 2024). D'altra parte, l'ASD non sindromico, noto anche come idiopatico, rappresenta la maggior parte dei casi e non è associato ad altre sindromi neurologiche note, sebbene coinvolga alterazioni nei geni correlati all'autismo (da Cruz Fontes e de Souza, 2022).
Le mutazioni genetiche ereditarie e de novo contribuiscono a fornire importanti spiegazioni per il disturbo. Le mutazioni ereditarie sono trasmesse dai genitori alla prole, mentre le mutazioni de novo si verificano nella linea germinale di una nuova generazione. È importante notare che le mutazioni non sono necessariamente dannose; esistono mutazioni neutre o benefiche, oltre a quelle che possono portare a disfunzioni biologiche, spesso associate a disturbi del neurosviluppo (Goldmann et al., 2019).
Mutazioni rare e ad alto impatto, in particolare quelle che colpiscono geni coinvolti nel rimodellamento della cromatina e nella segnalazione cellulare, sono state riconosciute come fattori critici nella predisposizione all'ASD. Questi cambiamenti genetici causano significative alterazioni morfologiche nel cervello, tra cui un aumento del volume cerebrale, in particolare nella corteccia prefrontale, anomalie nella densità e morfologia delle spine dendritiche, disfunzioni nelle reti di connettività funzionale e alterazioni nell'organizzazione della materia bianca e grigia. Tali disruzioni strutturali contribuiscono alla complessità fenotipica dell'ASD, riflessa nell'ampia variabilità dei deficit sociali, linguistici e comportamentali osservati tra gli individui affetti (Lim et al., 2022).
Le prove emergenti indicano che la disfunzione epigenetica, che coinvolge la metilazione del DNA e la deacetilazione degli istoni, contribuisce alla patogenesi dell'ASD, in particolare quando modulata da fattori di rischio ambientali durante la gestazione. Questi meccanismi epigenetici, essenziali per la regolazione dell'espressione genica, controllano l'attività trascrizionale: la metilazione nelle isole CpG reprime la trascrizione in condizioni di ipermetilazione e la favorisce in condizioni di ipometilazione. La disregolazione di questi processi può comportare l'attivazione o l'inibizione inappropriata di geni cruciali per lo sviluppo neurale, influenzando la funzione cerebrale sia a livello strutturale che funzionale (Genovese e Butler, 2023).
La deacetilazione degli istoni, promossa dalle istone deacetilasi (HDAC), induce la compattazione della cromatina e riduce l'accessibilità del DNA, con conseguente compromissione della trascrizione di geni cruciali per la formazione e il mantenimento delle sinapsi eccitatorie glutamatergiche e inibitorie GABAergiche. Di conseguenza, le alterazioni in questi processi influenzano le regioni cerebrali centrali per la fisiopatologia dell'ASD, come il giro cingolato, l'amigdala, lo striato (caudato-putamen) e il cervelletto, compromettendo i circuiti neurali associati alla cognizione sociale, alla regolazione emotiva, al controllo motorio e allo sviluppo del linguaggio (Torres et al., 2023).
Inoltre, le variazioni del numero di copie (CNV) rappresentano un importante meccanismo genetico associato alla patologia dell'ASD. Queste alterazioni, che coinvolgono delezioni o duplicazioni di segmenti di DNA, sono le mutazioni osservate più frequentemente e possono essere ricorrenti o non ricorrenti. Se localizzate in regioni critiche, come il locus 16p11.2, le CNV sono spesso correlate all'ASD, impattando significativamente il funzionamento biologico (Zhang et al., 2024).
Oltre alle variazioni CNV, si stima che almeno il 5% dei casi di ASD sia correlato a polimorfismi a singolo nucleotide (SNP) in geni come NLGN3, NLGN4, NRXN1, MECP2, SHANK3, FMR1, TSC1/2 e UBE3A. Circa il 10% delle CNV identificate colpisce direttamente la codifica proteica attraverso duplicazioni o delezioni geniche (Jiang et al., 2022).
La complessità dell'ASD, così come le mutazioni ad esso associate, non è dovuta all'azione isolata di specifiche alterazioni genetiche, ma piuttosto alla convergenza di queste mutazioni in comuni vie cellulari e molecolari. Queste vie sono particolarmente correlate a processi fondamentali come la trascrizione e la traduzione genica, i meccanismi epigenetici, le risposte immunitarie e infiammatorie, nonché la formazione e la funzionalità delle sinapsi (Jiang et al., 2022).
La formazione e la maturazione delle sinapsi sono essenziali per lo sviluppo dei circuiti neurali, con la disfunzione sinaptica come potenziale fattore patogeno coinvolto nei disturbi del neurosviluppo. Pertanto, il ruolo dei geni coinvolti in questa funzione ha guadagnato una significativa attenzione, comprese le mutazioni in geni come MECP2, SHANK, FMR1 (Zhang et al., 2024).
Il gene FMR1, la cui mutazione è causalmente associata alla sindrome dell'X fragile, è considerato anche la principale causa genetica nota di autismo. Questa sindrome è causata dal silenziamento del gene FMR1 a causa dell'ipermetilazione di una sequenza ripetuta di trinucleotidi CGG situata nella regione promotrice del gene, impedendone la normale espressione. La proteina FMRP, cruciale per il corretto funzionamento neuronale, si lega all'RNA e svolge ruoli chiave nella regolazione della traduzione proteica nei neuroni, nella stabilità dell'RNA, nella risposta ai danni del DNA e nella modulazione dei canali ionici, processi essenziali per l'omeostasi e la comunicazione neuronale (Milla et al., 2023).
Il gene MECP2, d'altra parte, codifica una proteina regolatrice trascrizionale essenziale per lo sviluppo e la funzione neuronale. Le mutazioni in questo gene sono la causa principale della sindrome di Rett, sebbene siano state associate anche all'autismo e all'encefalopatia neonatale (Gonzales e LaSalle, 2010). Espressa nelle cellule staminali neurali e gliali, MECP2 ha un'espressione più elevata nei neuroni post-mitotici. La sua funzione principale è quella di modulare l'espressione genica regolando la struttura della cromatina, agendo sia come repressore che come attivatore, a seconda del contesto cellulare. Le mutazioni più critiche si verificano nei domini MBD e NID, compromettendo l'interazione con il complesso NCOR1/SMRT e compromettendo la formazione di condensati nucleari, meccanismi chiave per la regolazione dell'attività genica nel b…
Autismo e genetica
Il disturbo dello spettro autistico (ASD) è una complessa condizione del neurosviluppo caratterizzata da sfide nella comunicazione, difficoltà nelle interazioni sociali e comportamenti ripetitivi che possono ostacolare lo sviluppo di un bambino. La crescente prevalenza dell'autismo richiede un'identificazione precoce e strategie di intervento efficaci. Questa revisione riassume le conoscenze attuali sui primi indicatori dell'ASD, inclusi i marcatori dello sviluppo cerebrale e i segni comportamentali visibili nei neonati. Indaga i processi diagnostici, sottolineando l'importanza di una diagnosi tempestiva tra i 18 e i 24 mesi utilizzando strumenti di screening consolidati. Discutiamo una varietà di approcci terapeutici, tra cui interventi comportamentali, strategie educative come la musicoterapia e progressi tecnologici come i dispositivi per la generazione del linguaggio. Inoltre, indaghiamo le opzioni farmacologiche per il trattamento dei sintomi associati, sottolineando la mancanza di farmaci mirati per i sintomi principali dell'ASD. Infine, presentiamo prove che evidenziano gli effetti positivi dell'intervento precoce sugli esiti dello sviluppo, sostenendo piani di trattamento individualizzati per migliorare il benessere dei bambini con ASD.
Introduzione
Il Disturbo dello Spettro Autistico (ASD) è una condizione eterogenea del neurosviluppo manifestata da difficoltà nella comunicazione, nell'interazione sociale e nell'engagement, accompagnate da comportamenti stereotipati, ripetitivi o interessi focalizzati che impattano varie aree dello sviluppo di un bambino [1]. Le stime sulla diffusione dell'ASD sono aumentate negli ultimi decenni, sottolineando l'importanza dell'identificazione precoce e della necessità di strategie di intervento efficaci [1]. Rimane incerto quanto l'aumento della prevalenza dell'ASD rifletta un vero aumento dei casi. Una maggiore identificazione e diagnosi, insieme all'espansione dei criteri diagnostici nelle varie edizioni del Diagnostic and Statistical Manual of Mental Disorders, sono probabili fattori che influenzano le stime di prevalenza [1,2]. Studi suggeriscono che alcuni bambini possano ricevere una diagnosi accurata di ASD già a due anni [3], sebbene casi più sottili potrebbero non manifestarsi fino a un'età più avanzata. Nonostante la maggiore consapevolezza dei primi segni, l'età media della diagnosi rimane tra i 4 e i 5 anni [4]. L'eziologia dell'ASD rimane elusiva, ma recenti scoperte suggeriscono una complessa interazione di cause genetiche, epigenetiche e ambientali [5]. I principali fattori di rischio includono il sesso maschile e una storia familiare di ASD, con una probabilità di ricorrenza nei fratelli minori che varia dal 7% al 19% [6,7], rispetto all'1,5% nella popolazione generale [2]. Il livello di rischio dipende dalla vicinanza delle relazioni familiari. Ad esempio, uno studio recente dalla Svezia ha riportato un aumento di dieci volte del rischio relativo se un fratello/sorella ha l'ASD, rispetto a un aumento di due volte se la condizione è presente in un cugino [8]. Meccanismi ambientali potenzialmente collegati all'ASD includono infiammazione, stress ossidativo e alterazioni ormonali [9,10].
Le evidenze supportano sempre più che una diagnosi precoce e interventi mirati possano avere un impatto positivo su vari domini dello sviluppo per i bambini con ASD [11]. La ricerca indica che la diagnosi precoce non solo migliora gli esiti dello sviluppo, ma anche l'integrazione sociale e il benessere generale a lungo termine [6]. Nonostante l'importanza della diagnosi precoce dell'ASD, le variazioni nelle procedure cliniche e la mancanza di strumenti diagnostici standardizzati portano a discrepanze nel riconoscimento, identificazione e trattamento precoci dell'ASD [12]. L'incoerenza nell'efficacia degli interventi terapeutici per l'ASD è dimostrata da alcuni studi che mostrano tassi di successo variabili e risultati distinti basati su fattori come condizioni comorbide e gravità dei sintomi [13,14]. Questi risultati incoerenti sottolineano l'importanza di studi futuri volti a migliorare i criteri diagnostici e le procedure di intervento, garantendo che i trattamenti siano tempestivi e personalizzati.
In questa revisione, presentiamo un riassunto dei predittori precoci dell'ASD, delle caratteristiche neuroevolutive e degli interventi. Mira a chiarire la complessità della diagnosi precoce, le variazioni neurobiologiche identificate nei neonati e l'impatto di varie tecniche terapeutiche. Combinando i risultati della ricerca esistente ed enfatizzando le lacune nella pratica clinica, questa revisione intende informare e guidare studi e trattamenti futuri, migliorando infine l'identificazione precoce e gli esiti del trattamento per i bambini diagnosticati con ASD.
Strategia di Ricerca, Identificazione degli Studi e Caratteristiche degli Studi
Questa revisione narrativa fornisce una panoramica della ricerca riguardante l'ASD nei bambini, concentrandosi specificamente sui primi segni e sugli interventi terapeutici. Per identificare studi pertinenti, sono stati utilizzati quattro database: Google Scholar, PubMed, Web of Science e Cochrane Library. È stata applicata la seguente sintassi booleana di ricerca: ((Autism Spectrum Disorder) OR (ASD)) OR (autism) AND ((((((“early signs”)) OR (“behavioral indicators”)) OR (“therapeutic interventions”)) OR (“intervention”)) OR (“early diagnosis”)) OR (screening). Successivamente, sono stati applicati vari filtri, tra cui: disponibilità del testo (testo completo), specie (umani), lingue (inglese) e un periodo di pubblicazione degli ultimi trent'anni. Infine, è stata effettuata un'ulteriore selezione utilizzando criteri di inclusione ed esclusione specifici, quali:
(i) Pubblicazioni in inglese;(ii) Studi pubblicati tra il 1990 e il 2024;(iii) Focus sui predittori precoci e sugli interventi terapeutici per l'ASD;(iv) Disegno dello studio: studi randomizzati controllati o revisioni sistematiche.
Gli studi sono stati esclusi se non soddisfacevano questi criteri o se mancava un focus diretto sui primi segni o sugli interventi terapeutici pertinenti all'ASD.
Sviluppo Cerebrale e Indicatori Precoci di ASD: Evidenze da Studi Longitudinali
Numerose ricerche longitudinali prospettiche che indagano potenziali biomarcatori basati sul cervello hanno rivelato varie differenze nello sviluppo neurale che appaiono intorno o prima della comparsa degli indicatori comportamentali precoci dell'ASD [12]. Diverse tecniche sono state utilizzate per valutare la struttura e la funzione del cervello nei neonati, ognuna offrendo una prospettiva distinta sullo sviluppo neurale. Le intuizioni dagli studi di RM, insieme ai primi risultati sull'autismo associato a sindromi utilizzando modelli animali, indicano variazioni nella crescita e nello sviluppo dei progenitori neurali e nella neurogenesi [13]. Cambiamenti nell'equilibrio tra processi inibitori ed eccitatori all'attività neuronale e sinaptica possono anche influenzare la formazione iniziale della circuitazione cerebrale funzionale nell'ASD, come indicato da una varietà di alterazioni elettroencefalografiche (EEG) osservate durante il primo anno di vita [14]. Mentre le caratteristiche comportamentali si sviluppano, i modi in cui i neonati interagiscono con il loro ambiente probabilmente influenzano ulteriormente la crescita della circuitazione neurale dipendente dall'esperienza.
Risultati della Risonanza Magnetica
Studi prospettici di risonanza magnetica (RM) hanno rivelato differenze significative nello sviluppo neurale precoce dei neonati che vengono successivamente diagnosticati con ASD. In particolare, un'eccessiva crescita cerebrale, caratterizzata da un aumento del volume della corteccia cerebrale, si verifica entro i primi due anni di vita tra i fratelli neonati che saranno identificati con ASD [15]. Questi neonati mostrano una rapida espansione della superficie corticale, in particolare nelle regioni occipitale, temporale e frontale, dai sei ai dodici mesi, che porta a un ulteriore sviluppo cerebrale eccessivo dai dodici mesi ai due anni [15]. Inoltre, viene osservata una crescita accelerata dell'amigdala nei neonati con fratelli diagnosticati con ASD a sei e dodici mesi, rispetto a quelli con sindrome dell'X fragile e sviluppo neurotipico [16].
Uno studio su cinquanta neonati ha mostrato una crescita nelle regioni sottocorticali tra i neonati di quattro-sei mesi, indipendentemente dalla diagnosi [17]. Un altro studio longitudinale ha rivelato che entro i dodici mesi, i neonati che sarebbero stati successivamente diagnosticati con ASD avevano strutture sottocorticali più grandi rispetto a quelli con ritardi del linguaggio precoci o senza autismo [18]. Inoltre, la ricerca che confronta 270 fratelli neonati con 108 neonati a bassa probabilità ha mostrato che un corpo calloso ingrossato a sei mesi prediceva futuri comportamenti correlati all'ASD, sebbene le variazioni nello spessore del corpo calloso tra bambini diagnosticati e non diagnosticati diminuissero al raggiungimento dei due anni, indicando un meccanismo di sviluppo dinamico [19].
Nella ricerca che coinvolge 92 fratelli neonati, le misurazioni dell'anisotropia frazionaria hanno indicato differenze nello sviluppo della materia bianca tra coloro che sono stati successivamente diagnosticati con ASD e coloro che non lo sono stati. I neonati con ASD hanno mostrato un'anisotropia frazionaria più elevata a 6 mesi, seguita da un cambiamento più graduale fino a ventiquattro mesi [20]. Allo stesso modo, uno studio su 116 fratelli neonati ha rivelato che quelli successivamente diagnosticati con ASD presentavano variazioni nell'efficienza della rete della materia bianca a 6 mesi [21]. Differenze nella maturazione della materia bianca in specifiche regioni cerebrali sono state collegate a particolari comportamenti correlati all'autismo [13].
Uno studio longitudinale che coinvolge 221 fratelli neonati ad alto rischio e 122 neonati a basso rischio ha rivelato che livelli elevati di liquido cerebrospinale extra-assiale (CSF) a 6 mesi predicevano l'ASD successivo, con livelli elevati persistenti fino a ventiquattro mesi [22]. Si ritiene che il CSF contenga fattori di crescita che supportano la proliferazione neuronale e possano aiutare a eliminare metaboliti che possono influenzare la funzione cerebrale.
Studi di risonanza magnetica funzionale a stato di riposo (fMRI) durante il sonno hanno identificato disturbi nella connettività talamocorticale e nelle reti legate al linguaggio già a 1,5 mesi nei fratelli neonati, indipendentemente dalla diagnosi futura [23]. A 6 mesi, pattern atipici di connettività funzionale nella fMRI erano collegati a una maggiore probabilità di identificazione dell'autismo entro i due anni [24].
Sebbene la maggior parte degli studi di RM si concentri sulle differenze di gruppo, alcuni hanno esaminato predizioni a livello individuale. Un algoritmo a 6 mesi che combinava CSF extra-assiale, volume cerebrale, età e sesso erano predittori della diagnosi di autismo con il 66% di sensibilità e il 68% di specificità [22]. Analisi di machine learning cross-validate di fMRI a stato di riposo alla stessa età hanno raggiunto l'82% di sensibilità e il 100% di specificità per la predizione dell'autismo [24]. Inoltre, le valutazioni dell'area della superficie corticale da sei a dodici mesi hanno predetto gli esiti con l'88% di sensibilità e il 95% di specificità [15].
Predittori Elettrofisiologici
Registrazioni della risposta uditiva del tronco encefalico, EEG spontanei e potenziali evocati (ERP) hanno dimostrato differenze in timing, ampiezza e potenza spettrale nei neonati che potrebbero fungere da predittori per l'ASD. Un esame delle precedenti risposte uditive del tronco encefalico da uno screening uditivo che ha coinvolto 139.154 neonati (321 successivamente diagnosticati con ASD) ha rivelato variazioni prolungate nella fase delle risposte uditive del tronco encefalico e una latenza della V-onda negativa osservate in coloro che sono stati diagnosticati con ASD [25]. Studi di machine learning su dati EEG spontanei da 99 fratelli neonati ad alto rischio e 89 neonati a basso rischio hanno indicato che la complessità dell'EEG e varie densità spettrali di potenza potevano predire l'autismo successivo [26]. Sebbene uno studio abbia collegato un'elevata connettività nella banda alfa dell'EEG a 14 mesi a una successiva diagnosi di autismo [27], un altro ha riscontrato che questa connettività era correlata solo a interessi limitati e comportamenti ripetitivi [28]. Tra i neonati di otto mesi, un'elevata reattività corticale ai toni ripetuti è stata associata a una successiva diagnosi di autismo, evidenziata da una ridotta soppressione delle risposte gamma a 40-60 Hz e da un'elevata coerenza inter-trial a 10-20 Hz [29]. A sei mesi, i neonati che sono stati successivamente diagnosticati con ASD hanno mostrato ERP più deboli in risposta alle espressioni facciali e tempi di attenzione visiva più brevi ai volti rispetto ai coetanei senza ASD [30]. Inoltre, i neonati di età compresa tra sei e dieci mesi hanno mostrato una ridotta coerenza inter-trial nella banda di frequenza theta durante l'elaborazione di stimoli visivi facciali [31]. I neonati diagnosticati con autismo a 8 mesi hanno mostrato una diminuzione dell'ERP N290, che è tipicamente reattivo agli stimoli facciali [32]. L'accuratezza predittiva per l'ASD successivo è migliorata quando i punteggi poligenici dell'autismo sono stati inclusi nel modello logistico che valuta le predizioni derivate dalle risposte di latenza N290 a stimoli facciali e non facciali [32].
In un piccolo studio randomizzato controllato (RCT) che ha coinvolto fratelli neonati di età compresa tra nove e undici mesi (n = 33), sono state utilizzate misurazioni EEG per valutare l'impatto di un intervento precoce volto a migliorare l'interazione sociale durante gli impegni caregiver-neonato. I neonati che hanno ricevuto questo intervento somministrato dal caregiver hanno mostrato pattern di sviluppo sia nell'EEG (potenza theta frontale) che negli ERP (risposta P400 ai volti) che si allineavano più strettamente a quelli dei neonati neurotipici, differendo da quelli che non hanno ricevuto l'intervento [33].
In un altro studio che ha coinvolto 91 fratelli neonati e 40 neonati a basso rischio di età pari a otto mesi, durate più brevi di microstati EEG associati all'attenzione sociale hanno predetto la futura diagnosi di autismo [34]. Inoltre, uno studio con 161 fratelli neonati ad alto rischio e 71 neonati a basso rischio ha utilizzato l'analisi delle componenti indipendenti per identificare pattern attraverso varie misure cognitive e adattive, comportamenti correlati all'ASD e risposte ERP a spostamenti dello sguardo, con l'obiettivo di scoprire pattern cross-dominio collegati a successive diagnosi di autismo [35].
La ricerca sulle predizioni a livello individuale indica che i biomarcatori elettrofisiologici possono essere efficaci per lo screening precoce dell'autismo, dimostrando alta sensibilità e specificità già entro i primi tre mesi di vita. In uno studio longitudinale che coinvolge fratelli e neonati a basso rischio, le traiettorie di potenza dell'EEG, in particolare nelle bande di frequenza delta e gamma, da tre a dodici mesi, hanno predetto accuratamente le successive diagnosi di ASD tra i fratelli neonati, raggiungendo una sensibilità di 0,82 e una specificità di 0,86 [26]. Inoltre, un algoritmo che incorpora caratteristiche EEG non lineari (come valutazioni di entropia e fluttuazione detrendizzata) è stato in grado di predire diagnosi di autismo in fratelli neonati a soli tre mesi, dimostrando una sensibilità di 0,82 e una specificità di 0,99 [36]. Un'analisi delle registrazioni cliniche della risposta uditiva del tronco encefalico da zero a tre mesi ha rilevato che una latenza estesa dell'onda V poteva differenziare tra trenta neonati successivamente diagnosticati con ASD e trenta neonati di controllo abbinati, con una sensibilità di 0,70 e una specificità di 0,80 [25].
Intuizioni dalla Spettroscopia nel Vicino Infrarosso
Sebbene la spettroscopia nel vicino infrarosso (NIRS) offra una risoluzione spaziale inferiore rispetto alla fMRI, è vantaggiosa per l'uso con neonati e bambini piccoli durante le attività [37]. In uno studio NIRS focalizzato su neonati di cinque mesi, comprendente 16 fratelli neonati e 13 neonati a basso rischio, i ricercatori hanno esaminato l'attività cerebrale in risposta a video sociali (come i comportamenti di un'attrice) e li hanno confrontati con le reazioni a stimoli non sociali (come immagini di automobili). I neonati a basso rischio hanno mostrato…
tags: #google #scholar #articoli #in #inglese #sull